
[transkribierter Text; kleinere Mängel waren wohl kaum zu vermeiden. Wer vollkommen exakte Daten und Fakten braucht, sei auf den Originaltext verwiesen. WM]
1. Einleitung
Im Spätsommer und Herbst streben alljährlich Millionen von Kleinvögeln — vorwiegend in südwestlicher Richtung — dem Mittelmeerraum zu. Nicht niedriger werdende Temperaturen, nicht die Veränderung des Nahrungsangebotes veranlassen die Vögel zum Zug. Die Zugauslösung erfolgt vielmehr im Rahmen eines angeborenen endogenen Jahresrhythmus, der vorwiegend fotoperiodisch, d. h. in Abhängigkeit von der wechselnden Tageslänge, gesteuert wird. Fast alle auf Insekten spezialisierte Arten verlassen im Herbst ihre europäischen Brutgebiete, während es unter den Früchte- und Körnerfressern Arten gibt, die sich dem wechselnden Nahrungsangebot angepasst haben und den Winter über im Brutgebiet bleiben. Die Masse der ziehenden Arten, die sogenannten Weitstreckenzieher (z. B. Gartengrasmücke, Fitis, Rauchschwalbe), überwintert südlich der Sahara. Zwischen diesen beiden Gruppen ordnet man die Kurzstreckenzieher (z. B. Rotkehlchen, Zilpzalp, Singdrossel, Hausrotschwanz) ein, die in der Regel nicht über den Mittelmeerraum hinausziehen (z. B. Zink 1973—1985, Hilgerloh 1987, 1989a).
Trotz dieses — schon allein zahlenmäßig — beeindruckenden und alljährlich. Wiederkehrenden Schauspiels sind zusammenfassende Darstellungen des Kleinvogelzugs aus dem europäischen Binnenland noch immer relativ selten (Ausnahmen: z. B. Dorka 1966, Berthold & Dorka 1969, Blondel 1969, Bairlein 1981, Brensing 1989). Für die britischen Inseln stellen Riddiford & Findley (1981) den Kleinvogelzug anhand von zahlreichen, anschaulichen Diagrammen einzelner Zugstationen dar. Während man in jeder „Avifauna” ausführliche Angaben zu den Brutvorkommen findet, sucht man meist vergeblich präzise Aussagen zum Durchzug von Vogelarten. Dies gilt auch für die beiden „Avifaunen” in Nordrhein-Westfalen (Peitzmeier 1969, Mildenberger 1984). Selbst die diversen Bände des „Handbuchs der Vögel Mitteleuropas” machen da bei den Kleinvögeln kaum eine Ausnahme (z. B. Glutz 1986, 1989). Was zum Durchzug zu sagen ist, erschöpft sich meist in verbalen Beschreibungen zu Zughöhepunkten und Zugzeiträumen (z. B. anhand von Erst- und Letztbeobachtungen). Systematische Untersuchungen zur Phänologie einzelner Kleinvogelarten aus Nordrhein-Westfalen sucht man meist ebenfalls vergeblich (Ausnahmen: Sartor 1984, Loske 1990 b). Etwas besser sieht es mittlerweile bei großen und visuell leicht erfassbaren Arten (z. B. Greifvögel, Limikolen, Wasservögel) aus (z. B. Stichmann et al. 1969, Harengerd et al. 1973, Ziegler 1981, Klaffke 1984, Knoblauch 1986, Schlüpmann 1986, OAG Münster 1989).
Wie dürftig unsere Kenntnisse über den zeitlichen und räumlichen Verlauf des Kleinvogelzugs in Nordrhein-Westfalen (und anderswo) sind, soll im Folgenden aufgezeigt werden. Darüber hinaus soll gezeigt werden, wie sinnvoll Untersuchungen zur Ökologie und Zugphysiologie von Kleinvogelarten in Rastgebieten sind. Anhand von ersten und vorläufigen Daten, die mit Hilfe eines standardisierten Fangprogramms gewonnen wurden, soll ferner aufgezeigt werden, auf welche Weise sich entsprechende Daten gewinnen lassen. Es liegt in der Natur der Sache, dass hier nur einige Aspekte zu diesem umfassenden Fragenkomplex aufgezeigt werden können.
2. Phänologische Aspekte
Ganz allgemein gilt: Während Kurzstreckenzieher später im Jahr wegziehen, setzt der Zug bei den Transsaharaziehern bereits ein, wenn die Nahrungssituation noch optimal ist. Obwohl letztlich die Nahrungsverknappung der Grund für den Zug ist, wirkt sie nicht direkt zugauslösend. Der Zug beginnt nach einer inneren Uhr, nach der auch der Zeitpunkt für Brut und Mauser festgelegt wird. Diese innere Uhr ist nicht sehr genau und wird in der Natur anhand der Tageslänge mit den Jahreszeiten synchronisiert.
Jeder, der in eine „Avifauna” oder ein Handbuch schaut, sollte nun dort eigentlich Auskünfte über die Phänologie (Beginn, Höhepunkt und Ende des Zugs) einzelner Kleinvogelarten finden. Leider ist dem nicht so. Anstelle von präzisen Werten oder anschaulichen Diagrammen findet man fast nur verbale Umschreibungen („die Masse ist bis Ende September durchgezogen”). Ein Grund hierfür ist sicherlich die Tatsache, dass das Zuggeschehen von Kleinvögeln für den Beobachter nur in sehr begrenztem Maße wahrnehmbar ist. Viele Arten ziehen zudem nachts oder vorwiegend in großer Höhe (z. B. Hilgerloh 1989b).
Die objektivste Methode zur Registrierung des Durchzugs von Kleinvögeln ist zweifellos ein standardisiertes Fangprogramm. Das wohl bekannteste Beispiel dafür ist das „Mettnau-Reit-Illmitz-Programm” (Berthold et al. 1986 a). Hier wurde über Jahre hinweg die gleiche Anzahl Netze an jeweils denselben Stellen und in derselben Höhe: aufgestellt.
Zusätzlich wurde die Gebüsch-Vegetation in regelmäßigen Abständen auf die Höhe zu Beginn des Fangprogramms zurückgeschnitten. Einflüsse auf die Fangmuster durch wechselnde Standorte sind damit ausgeschlossen.
Die Zahlen von Durchzüglern schwanken häufig tageszeitlich, jahreszeitlich, witterungsbedingt und von Jahr zu Jahr stark. Zur Charakterisierung der Häufigkeitsverteilung von Fangdaten gibt es verschiedene Möglichkeiten. Ziel der Auswertung ist es in allen Fällen, die in den Zahlenreihen versteckten, zufälligen Einflüssen unterworfenen „Gesetzmäßigkeiten offenzulegen. Sehr anschaulich sind in jedem Fall Durchzugsdiagramme.
Welche Form man wählt (z. B. Tagessäulen, Pentaden-, Wochen- oder Dekadensäulen, Liniendiagramme), muss genau bedacht werden. Ausführliche Angaben zu den Vor- und Nachteilen dieser Darstellungsformen finden sich in der Literatur (z. B. Scharlau 1968, Gnielka 1972, Harengerd et al. 1973, Berthold et al. 1980).
Ein bei allen Untersuchungen auftretendes Problem ist die wechselnde Intensität des Fangaufwands. Die Berechnung zugphänologischer Kennziffern (s. u.) und/oder die Darstellung durchschnittlicher Tageswerte ist natürlich nur sinnvoll, wenn möglichst regelmäßig und vollständig gefangen wird, d. h. größere Erfassungslücken vermieden werden. Andernfalls muss eine Zusammenfassung zu Zeiträumen erfolgen. Eine sehr sinnvolle Darstellungsform wird z. B. bei Ormerod (1990) vorgestellt: Dort werden die Fangzahlen zur Vermeidung von Einflüssen durch wechselnde Fangintensität als „Anzahl der Expl. pro Fangstunde” (catch per hour) dargestellt. Selbstverständlich kann man auch andere Zeitabschnitte wählen.
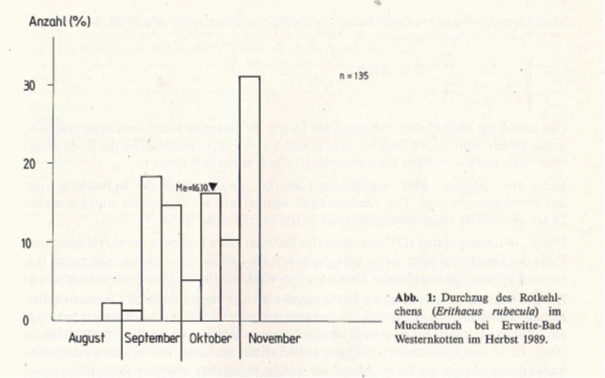
Als Beispiel für ein Diagramm auf Dekadenbasis zeigt Abb. 1 den Durchzug des Rotkehlchens (Erithacus rubecula) während der Herbstzugperiode 1989 im Muckenbruch bei Erwitte-Bad Westernkotten (vgl. Tab. 1). Unschwer sind drei „Durchzugswellen” (1. Mitte-Ende September; 2. Mitte Oktober; 3. Anfang November) zu erkennen. Der starke Durchzug Anfang November deutet darauf hin, dass auch noch nach dem 10. 11. Durchzug auftritt und der Median damit in Wirklichkeit später liegt.
Will man den „eigenen” Durchzug von Arten mit denen anderer Autoren vergleichen, sollte man auf jeden Fall: drei Kennziffern angeben: den Mittleren Zugtag (Mr), den Median (Me) und die Durchzugsdauer (D).
Der Mittlere Zugtag wird nach der Formel für das arithmetische Mittel berechnet:
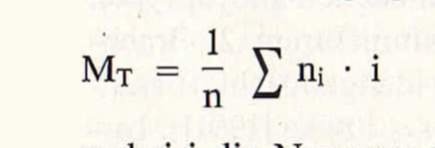
wobei i die Nummer des einzelnen Tages in einer Zugperiode angibt und ni; die Zahl der an diesem i-ten Tag registrierten Ex.; n ist die Summe aller n.
Als Mittelwert aller Durchzugsdaten kennzeichnet der Mittlere Zugtag im Allgemeinen den Höhepunkt des Zuges. Bei symmetrischem Verlauf des Durchzugsdiagramms sind bis zu diesem Tag 50 % der Durchzügler einer Art über das Gebiet hinweggewandert (Gnielka 1972). Bei Arten, die zur Überwinterung neigen oder im Gebiet brüten, lässt sich der Mittlere Zugtag nicht ohne weiteres berechnen, da ihn extrem frühe und späte Daten nachhaltig beeinflussen.
Man kann die Standardabweichung Mittleren Zugtag MT wie folgt berechnen:
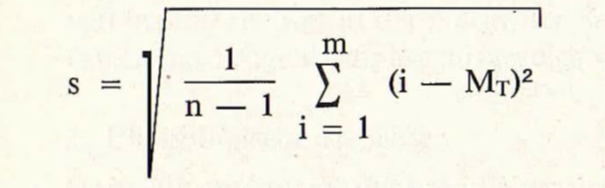
s ist damit ein Maß für die Streuung der Daten. Je länger sich der Durchzug hinzieht, umso größer wird in der Regel s. Im Bereich von Mr —s ziehen 68 % der Individuen einer Art durch — sofern die Zahlenreihen normalverteilt sind.
Unter dem „Median (Me)” versteht man den Tag, an dem 50 % der Individuen einer Art durchgezogen sind. Der Median wird weniger als der Mittelwert durch extreme Daten beeinflusst (Berechnungsbeispiel s. Harengerd et al. 1973).
Unter „Durchzugsdauer (D)” gibt man (im Rahmen eines Fangprogramms) Beginn und Ende des Durchzugs einer Art an. Zu bedenken ist, dass sich beide kaum abgrenzen lassen und extrem frühe und späte Daten die Aussagekraft dieses Zeitraums abschwächen.
Einige Autoren (z. B. Niemeyer 1975) empfehlen die Berechnung des geometrischen Mittels, die aber recht aufwendig ist und sich deshalb auch nicht durchgesetzt hat. Das am häufigsten angewendete Mittelwertmaß ist mittlerweile der Median (z. B. Bairlein 1981). Er ist von Extremwerten nahezu unbeeinflusst und zeigt im Vergleich zum arithmetischen und geometrischen Mittel die geringste mittlere absolute Abweichung von den Einzelwerten bei zugphänologischen Auswertungen (Klaffke 1987). Gleichwohl decken sich Median und Mittlerer Zugtag bei symmetrischem Verlauf der Durchzugskurve.
Leider hat sich bisher bei Kleinvögeln kaum jemand die Mühe gemacht, Diagrammdarstellungen oder Mittelwertmaße vorzulegen. Bedauerlicherweise lassen sich deshalb bisher kaum Vergleiche des Durchzugs zwischen verschiedenen Regionen ziehen. Aus Nordrhein-Westfalen sind mir überhaupt keine Publikationen bekannt, die die Zugphänologie von Kleinvögeln entsprechend darstellen. Gerade bezüglich der Phänologie der Arten ist derzeit also noch alles offen und entsprechende Untersuchungen sind sehr lohnenswert.
Unter anderem aus diesem Grunde habe ich 1989 im Muckenbruch bei Erwitte-Bad Westernkotten, Kreis Soest (51° 39” N, 8° 22’ E) versuchsweise eine Dauerfanganlage betrieben. Sie bestand aus insgesamt 69 m Netz, die in zwei verschiedenen Biotoptypen (1. Schilf-Weidengebüsch und Feuchtbrache mit Arten der Schlammfluren; 2. Brachacker mit Zaunwinden-Schleiergesellschaft und einzelnen Ohrweidengebüschen) standen (zur Lage, Charakterisierung und Brutvogelfauna des Gebietes s. Loske [1991]). Insgesamt erfolgten dort 61 jeweils mehrstündige Fangeinsätze zwischen dem 11. 8 und 10. 11. 1989. Dabei wurden 27 Kleinvogelarten in 1460 Individuen gefangen und beringt.
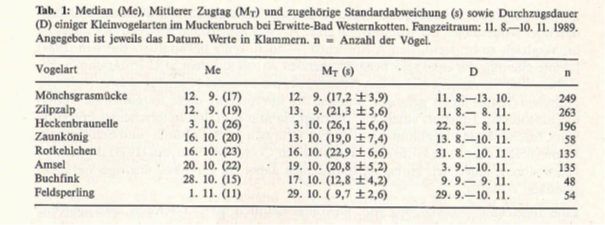
Zu danken habe ich Bernd Pohl (Erwitte), der für mich den Großteil der Fangarbeiten durchführte und Thomas Laumeier (Geseke), der mir bei der Auswertung half. Tab. 1 zeigt die Zugmediane, den Mittleren Zugtag (inkl. Standardabweichung) und die Durch zugsdauer für die Arten mit ausreichender Fangzahl. Soweit mir bekannt, werden damit für Westfalen erstmals Werte für den Durchzug dieser Arten vorgelegt. Ich hoffe, dass sie zu zukünftigen Vergleichen reizen.
Mit Ausnahme des Buchfinken liegen Median und Mittlerer Zugtag dicht beieinander. Hervorzuheben ist, dass der Beginn des Fanges am 11. 8. mit Sicherheit zu spät liegt — insbesondere für frühe Transsaharazieher, die in Tab. 1 nicht vertreten sind (Ausnahme: Mönchsgrasmücke). Laut MRI-Programm sollten die Fanganlagen vom 30. 6.—86. 11. betrieben werden. Nur dann dürften vor allem für frühe Arten wie z. B. Nachtigall, Feldschwirl und Gelbspötter realistische Mittelwerte zu erwarten sein.
Einschränkend ist nochmals deutlich hervorzuheben, dass die angegebenen Werte lediglich aus einer einzigen Zugperiode stammen. Um aber einen für einen bestimmten Raum repräsentativen Mittleren Zugtag zu bestimmen, sind die Daten vieler Jahre notwendig.
Ziel sollte es sein, eines Tages für jede ziehende Kleinvogelart eine Karte (z. B. im Rahmen eines Zugatlanten) vorlegen zu können, in der das Fortschreiten des Zuges in unterschiedlichen geographischen Räumen, z. B. anhand der Mediane, dargestellt wird. Für Limikolen existiert bereits ein Versuch, die zeitliche Lage des Wegzugs im west- und mitteleuropäischen Raum anhand von Zugparametern einheitlich darzustellen (OAG Münster 1988).
3. Ökologische Aspekte
Wir leben im Zeitalter der Renaturierungen und des Biotopmanagements. Kaum eine Ökologische Planung läuft heute noch ab, ohne dass — neben der obligatorischen Erfassung der Vegetation — wenigstens einige Tiergruppen untersucht werden. Diese Erfassung der Tierwelt gehört demnach ganz eindeutig zum Aufgabenkatalog ökologischer Grundinformationen (z. B. Blab 1978, 1986). Ohne diese ausführlichen Grundinformationen verkümmern geplante ökologische Verbesserungen schnell zur Garten- und Landschaftskosmetik. Die Erfassung der Brutvögel als Indikatoren für die Beurteilung einer Umweltsituation (s. Bezzel & Ranftl 1974, Utschik 1984) ist mittlerweile erfreulicherweise Standard in der Planungspraxis, durchziehende Kleinvögel spielen dabei aber offensichtlich gar keine Rolle. Allenfalls Wasser- und Watvögel scheinen dem Landschaftsplaner in diesem Zusammenhang mitunter einer Erwähnung und Beachtung wert. Bedenkt man, dass viele Vogelarten den größten Teil ihres Lebens auf dem Zug verbringen, dann muss die „Rastfunktion” vieler Gebiete bei ökologischen Planungen endlich ausreichend gewürdigt werden.
Die andauernde und konsequente Ignorierung der Durchzügler mag auch daran liegen, dass über Änderungen der Habitatwahl einer Art in Rastgebieten während des Zuges — im Vergleich zu brutzeitlichen Ansprüchen — noch wenig bekannt ist. Bairlein (1981) konnte allerdings in einer sehr beeindruckenden Arbeit aus dem MRI-Programm nachweisen, dass die meisten ziehenden Kleinvogelarten über Jahre immer wieder in für sie typischen Teilhabitaten gefangen wurden. Er interpretiert diese festgestellten Verhaltungsmuster als Ergebnis einer sehr präzisen Habitatwahl und unterscheidet mit Hilfe von Clusteranalysen bestimmte ökologische Gruppen (z. B. Gebüsch- und Schilfvögel). Ferner berechnet er mit Hilfe einer Formel nach Colwell & Futuyma (1971) die relative Nischenbreite einer Art. Hohe Werte bedeuten dabei ein euryökes, niedrige Werte ein stenökes Verhalten.
Eine allgemeine Einschätzung wird allerdings dadurch weiter erschwert, dass ziehende Kleinvögel in verschiedenen Gebieten auch unterschiedliche Habitate bevorzugen können. Zudem zeigen insbesondere spät ziehende Arten eines Gebietes ein — dem Nahrungsangebot angepasstes — Shiften der Biotopwahl, das auch ganz allgemein tages- und jahreszeitlich bedingt auftritt (Bairlein 1981, Brensing 1989). Wie eng wiederum phänologische und ökologische Aspekte miteinander verwoben sind, zeigt der Umstand, dass Arten gleicher Gattungen (die miteinander um Ressourcen konkurrieren können wie z. B. Zilpzalp und Fitis) in den Rastgebieten in aller Regel zeitlich getrennt durchziehen.
Für die arttypischen Verteilungsmuster (und damit für die Trennung ziehender Kleinvögel in Rastgebieten) dürften neben Mechanismen der (angeborenen) artspezifischen Habitatwahl, dem Nahrungsangebot und der Konkurrenz mit anderen Arten wohl vor allem morphologische Aspekte eine wichtige Rolle spielen. Die Morphologie einer Vogelart kommt dabei vor allem in den Funktionskreisen Nahrungserwerb und Lokomotion zum Tragen (Bairlein et al. 1986).
Leider scheint es über Bairlein (1981) hinaus keinerlei Vergleichsarbeiten zur Habitatselektion von Kleinvögeln in Rastgebieten zu geben — schon gar nicht aus Nordrhein-Westfalen. Ohne Kenntnis der ökologischen Ansprüche der einzelnen Arten (wobei aus Gesichtspunkten des Artenschutzes insbesondere gefährdete Arten von Interesse sind) lässt sich für diese Arten natürlich auch keine gezielte Schutzstrategie entwickeln. In Anbetracht der augenblicklich in NW hochaktuellen Naturschutzdebatten (Feuchtwiesen- und Auenschutz, Biotopmanagement für eine immer größer werdende Zahl von Schutzgebieten) ist dieser Zustand höchst bedauerlich. Auch Untersuchungen zur Biotop- und Vertikalverbreitung ziehender Kleinvögel mit Hilfe eines standardisierten Fangprogramms sind daher in höchstem Maße wünschenswert. Fernziel sollte dabei sein (soweit möglich), allgemeine Regeln für Habitatpräferenzen und ökologische Sonderungen von rastenden Zugvögeln zu finden (Berthold et al. 1986 a).
4. Zugphysiologische Aspekte / Mauser
Für ziehende Vogelarten spielt die Anlage von Fettdepots als Energievorrat eine bedeutende Rolle. Das Fett kann bis zu 100 % des ursprünglichen Körpergewichts betragen und wird überwiegend subkutan angelagert. Es ist am lebenden Vogel an dessen Färbung zu erkennen, weshalb der entsprechende Vogel skalierten Fettklassen zugeordnet werden kann (z. B. Pettersson & Hasselquist 1985). Diese Fettdepots liefern den Brennstoff für den aktiven Zug.
Da Fett als Flugtreibstoff fungiert, wird die Fettdeposition vor allem als eine Anpassung an die Überquerung großer ökologischer Barrieren (Mittelmeer, Sahara) aufgefasst. Die Fettanlagerung ist einem endogen kontrollierten jährlichen Rhythmus unterworfen, der sich auch unter Laborbedingungen nachweisen lässt (Übersicht s. Berthold 1975). Zugvögel — vor allem unerfahrene Jungvögel — verfügen über ein angeborenes Zugzeit-Programm, das ihnen den richtigen Zeitpunkt zum Aufbruch, die Zugweglänge und auch die Richtung für ihren Zug weitgehend vorgibt. So weist z. B. die in Laborversuchen an der Gartengrasmücke registrierte nächtliche Zugaktivität (Zugunruhe) dann ihr Maximum auf, wenn im Freiland die Überquerung von Mittelmeer und Sahara angenommen wird. Zudem ist die im Käfig produzierte Menge an Zugunruhe mit der Zugweglänge im Freiland positiv korreliert. Neueren Untersuchungen aus der Zentralsahara zufolge (z. B. Bairlein 1985a, 1987, Biebach 1986) scheint ein solches endogenes Programm im Freiland aber nicht sehr streng abzulaufen. So durchqueren die europäischen Kleinvögel die Sahara offensichtlich mehr in Etappen als in einem langen Nonstopflug (s. aber Wood 1989). Reicht der Fettvorrat nach einer Landung in einem Sahara-Rastgebiet noch für den Weiterzug aus, rasten die gut genährten Tiere inaktiv im Schatten, um in der nächstfolgenden Nacht weiterzuziehen. Magere Tiere hingegen müssen ihre Fettvorräte auffrischen und sind gezwungen, tagsüber auf Nahrungssuche zu gehen.
Auch für das mitteleuropäische Binnenland dürfen wir — zumindest für den Herbstzug — annehmen, dass die meisten Kleinvögel mehr schrittweise ziehen. Dies unterstreicht einmal mehr, welche wichtige Rolle Rastgebiete in der Biologie von Zugvögeln spielen. Offenbar werden hier die lebenswichtigen Fettvorräte für den Weiterzug angelegt, d. h. Zugunterbrechung erfolgt vor allem dann, wenn das Körpergewicht erniedrigt ist, die Fettreserven aufgebraucht sind und das Nahrungsangebot so gut ist, dass Fettvorräte aufgebaut werden können.
Im Rahmen eines standardisierten Fangprogrammes lassen sich auch solche Daten zur Zugphysiologie einfach gewinnen. Fang und Beringung erlauben Untersuchungen des Körpergewichts und der vorhandenen Fettdepots. Für die Ermittlung der Fettdepots gibt es mittlerweile eine Fülle von Methoden. Meines Erachtens ist die von Pettersson & Hasselquist (1985, mit Abbildungen) vorgestellte Methode am geeignetsten. Sie basiert auf insgesamt sieben Fettklassen (0 = kein Fett; 6 = der gesamte Bauch mit einer dicken Fettschicht überzogen), die jeweils geschätzt werden müssen. Von besonderem Interesse sind dabei Aussagen zur Verweildauer von Individuen, die sich über Wiederfänge gewinnen lassen. So sollten z. B. vorhandene Fettdepots und Aufenthaltsdauern negativ miteinander korreliert sein. Ferner lassen sich mit Hilfe der Fettdepots sogenannte „flight-ranges” berechnen (z. B. Davidson 1984). Dabei wird ermittelt, wie weit ein Vogel mit einem bestimmten Fettvorrat theoretisch fliegen kann, ohne landen zu müssen. Immerhin waren etwa 50 % der Rotkehlchen und Wintergoldhähnchen in Südschweden in der Lage, mit ihrem Fettvorrat mehr als zehn Stunden ununterbrochen zu ziehen. Dabei sollen 0,7 % (Wintergoldhähnchen) bzw. 0,9 % (Rotkehlchen) des Körpergewichts pro Flugstunde verbraucht werden. 1—5 % der untersuchten Vögel schafften es sogar, länger als 25 Stunden ununterbrochen zu fliegen. Wintergoldhähnchen benötigten danach vier Tage, um ihr Ausgangsgewicht wieder zu erreichen. (Pettersson & Hasselquist 1985).
Als Beispiel einer graphischen Darstellung der jahreszeitlichen Variation des Körpergewichts einer Kleinvogelart zeigt Abb. 2 den Verlauf des Körpergewichts männlicher Rauchschwalben (Hirundo rustica; entnommen aus Loske 1990). Auch wenn diese Daten nicht aus einem standardisierten Fangprogramm stammen, so zeigen sie doch, welche Möglichkeiten der Darstellung man mit Gewichtsreihen einer Kleinvogelart hat. Es ist zu erkennen, dass männliche Rauchschwalben im Frühjahr mit hohen Gewichten im Brutgebiet ankommen und kurz vor dem Wegzug wiederum stark an Gewicht zulegen. Auch ohne die tatsächliche Ermittlung von Fettklassen deutet dies darauf hin, dass Rauchschwalben im Frühjahr mit Fettdepots ankommen und solche vor dem Wegzug erneut anlegen.
Der Vollständigkeit halber sei hier noch auf einen weiteren, wichtigen Aspekt hingewiesen: Unsere Kenntnisse über die Mauser zahlreicher Kleinvogelarten sind — wohlwollend formuliert — äußerst dürftig (Übersicht bei Kasparek 1981). Standardisierte Fangprogramme bieten auch diesbezüglich gute Möglichkeiten. So lassen sich zum einen (über Wiederfänge) Daten zur Gefiederentwicklung einzelner Individuen sammeln. Ferner lassen sich für die in einem bestimmten Zeitraum untersuchte Population Mauserstadien (z. B. ausgedrückt in % der erneuerten Federn) für einzelne Gefiederpartien (z. B. Handschwingen) ermitteln (Berechnungsbeispiele in Loske & Lederer 1988). Da zahlreiche Kleinvogelarten ihre Vollmauser im Winterquartier durchmachen, kommen hier natürlich nur bestimmte Arten in Betracht.
5. Bestandsentwicklung
Alle hier angesprochenen Fragen stehen in engem Zusammenhang mit dem vieldiskutierten Rückgang ziehender Singvogelarten und der Suche nach den Ursachen dafür (Bairlein 1985b). Wie stark selbst Allerwelts-Arten, die man ja noch „überall” sieht, abgenommen haben, ist spätestens seit den dramatischen Ergebnissen des MRI-Programms bekannt (Berthold et al. 1986 b).
Eine über Jahre standardisiert betriebene Dauerfanganlage dient also dem gleichen Ziel wie eine Untersuchung zur Bestandsentwicklung von Brutvögeln. Bedenkt man, dass nach der Methode der Papierreviere (Berthold et al. 1980) mindestens zehn Begehungen pro Jahr zur Bestandsermittlung erforderlich sind, dann ergeben sich nach den Siedlungsdichte-Richtlinien (8 min/ha) bei einer 30 ha großen Fläche schon 40 h Kartierungszeit. Damit ist der Aufwand bei einer Dauerfanganlage so viel größer gar nicht. Auch hier dürften sich (eine ausreichende Anzahl Netze vorausgesetzt) schon ab etwa 15—20 Fangeinsätzen pro Jahr in einer Zugperiode (Mitte Juli—Ende Oktober) durchaus interessante Ergebnisse ermitteln lassen. Hinzu kommt, dass Brutvogel-Bestandsaufnahmen meist nur einen kleinen Ausschnitt einer Population beschreiben, der keineswegs repräsentativ sein muss. Um großräumige Populationstrends aufzuzeigen, sind Daten von Zugstationen möglicherweise zuverlässiger, da sie das Populationsgeschehen eines größeren Hinterlandes widerspiegeln (Gatter 1978). – In Großbritannien gibt es z. B. die sogenannten „Constant effort sites”, in denen alljährlich systematisch gefangen wird. Dieses Programm des British Trust for Ornithology (BTO) hat neben dem bekannteren „Common Bird Census” bereits sehr interessante Aufschlüsse über Bestandsentwicklungen geliefert (z. B. Hale 1986).
Zusammenfassung
In der vorliegenden Arbeit wird das derzeitige Wissen über phänologische, ökologische und physiologische Aspekte des Kleinvogelzugs in knapper Form resümiert — insbesondere mit Blick auf die Verhältnisse in Nordrhein-Westfalen. Im Muckenbruch-Niedermoor bei Erwitte-Bad Westernkotten (51° 39” N, 8° 22’ E, Kr. Soest), einem sehr heterogenen Biotopkomplex (Gehölze, Acker, Grünland, Bachlauf, Schilf, Moorbrachen) wurde vom 11. 8.—6. 11. 1990 eine Dauerfanganlage mit 69 m Netz betrieben. Insgesamt wurden dort 1460 Kleinvögel in 27 Arten gefangen. Für acht Arten werden erstmals vorläufige, zugphänologische Kennziffern (Median, Mittlerer Zugtag und Standardabweichung, Durchzugsdauer) vorgelegt, die zu Vergleichsuntersuchungen anregen sollen. Es wird betont, dass auch Daten zur Habitatselektion, Zugphysiologie und Mauser von Kleinvögeln Mangelware sind. Der Betrieb von standardisierten Dauerfanganlagen bietet gute Möglichkeiten, unsere diesbezüglichen dürftigen Kenntnisse zu ergänzen.



Anschrift des Verfassers: In den Kühlen 44, 4787 Geseke